 Sciencemag.cz
Sciencemag.cz

Related Articles


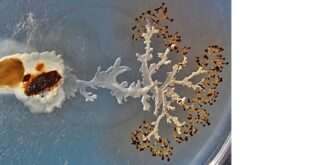
Rybomorky, buňky HeLa a jednobuněčný pes. Nové tělní plány rozhodně nevznikaly jen v kambriu.
Podívejme se na několik zvláště působivých případů, kde relativně nedávný (obvykle pozdně prvohorní či druhohorní, někdy i mnohem pozdější, ale rozhodně ne kambrijský) vznik absolutně nových tělních plánů v rámci „kmenů“ těžko zpochybnit.
Kořenohlavci (Rhizocephala) jsou parazitičtí korýši obývající těla jiných korýšů, například krabů. Ačkoli jsou parazitická zvířata často velmi podivná, podivnost kořenohlavců přece jen překračuje všechny meze. Z vajíčka se líhnou planktonní larvy obvyklého korýšího typu (podle larev jsme také už roku 1787 poznali, že to jsou korýši). Larva usedne na svého budoucího hostitele a vytvoří stadium kentrogon s jakousi injekci, jíž vtlačí dovnitř hostitele červovité stadium vermigon. Vermigon se postupně začíná větvit – a vytváří tak složitou síť trubiček vysávajících hostitele, ne nepodobnou myceliu hub. Na této síti pak vznikne vakovitý útvar, který po dalším svlékání hostitele vyhřezne na povrch těla. Toto je vývojový cyklus samice kořenohlavce; samec rovněž začíná larválním stadiem, ale poslední larva přisedne na vyhřezlou parazitickou samici a injikuje další stadium, trichogon, nikoli do hostitele, nýbrž do samice. Tam dojde k oplození a z oplozených vajíček se líhnou larvičky. Vermigon a trichogon jsou extrémně zjednodušená stadia členovců omezená na pouhou pokožku, kutikulu a shluk buněk, z nichž se postupně vytvářejí pohlavní žlázy. Nicméně nelze hovořit o pouhém morfologickém zjednodušení; zatímco vermigon můžeme jakžtakž považovat za korýše bez hlavy, bez nohou, bez vnitřních orgánů a dokonce i bez nervové soustavy, vlastní rozvětvené parazitické stadium je strukturálně docela složité, ale absolutně nekorýší.
Rybomorky (Myxosporea) najdete ve starších učebnicích mezi parazitickými prvoky. Jsou to nicméně „prvoci“ zvláštní: vlastního parazita v těle ryby představuje mnohojaderný útvar, v němž vznikají vícebuněčná infekční stadia, spory. Struktura buněk spor i jejich ontogeneze naznačovaly už v „době předmolekulární“, že jde o mnohobuněčné živočichy extrémně pozměněné parazitismem a že nejbližší příbuzné rybomorek můžeme hledat mezi žahavci (tedy nezmary či medúzami). „Žahavčí hypotéza“ je zdaleka nejpravděpodobnější i po přečtení genomů rybomorek; přinejmenším si můžeme být absolutně jisti, že jde skutečně o zjednodušená mnohobuněčná zvířata pocházející z vysoce organizovaných předků, kteří už měli nervovou i svalovou tkáň. Redukce na několik málo (4–6) buněk je zcela bezprecedentní ztrátou většiny živočišných vlastností.
A konečně proč chodit tak daleko. V roce 1951 byla v americkém Baltimoru jednatřicetileté Henriettě Lacksové diagnostikována rakovina děložního hrdla a několik buněk bylo odesláno k laboratornímu vyšetření. Sama Henrietta zemřela o osm měsíců později, ale potomci jejích rakovinných buněk, takzvané buňky HeLa, se dosud kultivují v laboratořích po celém světě, a dokonce i létají na oběžnou dráhu. Kolonie buněk HeLa evidentně není člověk, má úplně jinou morfologii, fyziologii (neboť jde o jednobuněčné organismy) i ekologii (žije v laboratorním skle), přesto ale má docela dobře rozpoznatelný hominoidní genom. Před očima nám vznikl jednobuněčný savec, jednobuněčný primát, jednobuněčný člověk, tedy dosti zřetelně nový „tělní plán“, a to nikoli v kambriu, nýbrž před šedesáti lety. Dobrá, to vypadá jako pouhá spekulace, která nemá s přírodou a evolucí nic společného.
Jenže nedávno se zjistilo, že psí přenosný pohlavní nádor (CTVT) není jen tak obyčejné nádorové onemocnění. Normální rakovinu tvoří prostě zdivočelé buňky, které nicméně i nadále zůstávají nedílnou součástí těla, kde vznikly, a nemohou je opustit; vznik rakoviny je nevyhnutelným důsledkem schopnosti buněk množit se a adaptovat. Právě proto, že jde o naše vlastní tělní buňky, náš imunitní systém je nedokáže odhalit a zlikvidovat; naopak transplantujeme-li nádor jinému jedinci, jeho imunitní systém se s ním vypořádat dokáže. Přesto jsou mnohé nádory „nakažlivé“; většinou to znamená, že nádor je vyvolán nakažlivým činitelem, obvykle virem. CTVT je ale něco úplně jiného. To, co se tu přenáší ze psa na psa, není činitel vyvolávající v novém hostiteli novou rakovinu, nýbrž klon starých rakovinných buněk jako takových. Rakovinné buňky z mnoha nepříbuzných psů z Evropy, Asie i Afriky jsou geneticky skoro totožné a fylogenetická analýza ukázala, že jde o potomky jedné jediné prabuňky (patrně myeloidní buňky z kostní dřeně), která se zvrhla před nějakými deseti tisíci let (u takto extrémních deviantů se stáří špatně odhaduje, tady molekulární hodiny tikají těžko odhadnutelnou rychlostí, ostatně přes 600 psích genů CTVT za tu dobu ztratil úplně). Můžeme mluvit o nejdéle existující kontinuální linii savčích somatických („tělních“) buněk, o jakési spontánně vzniklé a spontánně se udržující „tkáňové kultuře“ (Murchison a spol., 2014). Anebo z druhé strany o novém, nepohlavně se množícím, jednobuněčném a parazitickém druhu psovité šelmy, která napadá jiné psovité šelmy (Frank, 2007). Savec tedy nemusí být jenom mnohobuněčný. Nejde jenom o psy, kde se původ klonálního rakovinného psa ztrácí v neurčité minulosti – infekční rakovina obličeje (DFTD), decimující populace tasmánských ďáblů medvědovitých (Sarcophilus harrisii), má stejnou povahu a vznikla zvrhnutím jedné nervové Schwannovy buňky v jedné samici před pár desetiletími (první záznam choroby pochází z roku 1996). DFTD se od CTVT liší především tím, že silně metastázuje a je většinou smrtelný. Proto se nezdá, že by s námi mohl být tak dlouho jako vcelku neškodný nádoropes; válka dvou ďáblů, dravého a parazitického, se už chýlí ke konci, a nezasáhne-li vyšší (zde ochranářská) moc, druhý porazí prvního a zanikne s ním (Siddle a Kaufman, 2013).
Těžko považovat kořenohlavce a rybomorky za pouhé „drobné modifikace“ členovčího a žahavčího tělního plánu, o jednobuněčné formě Henrietty Lacksové a nakažlivých ďáblech ani nemluvě. Můžeme si připravit zadní vrátka a prohlásit, že jde vlastně vždycky o druhotné redukce již hotových tělních plánů, že to není evoluce, ale jen „devoluce“. Jenže tím si nepomůžeme: strukturální zjednodušení nemusí být evolučně nijak snadné. Rybomorka žije bez orgánů i bez tkání, ale žití bez orgánů a tkání vyžaduje jakýsi biologický grif, který my ostatní neovládáme; také žije v úplně jiných podmínkách než její volně žijící žahavčí předkové a na tyto podmínky se musela dlouhodobě adaptovat.
Navíc nemusí jít vždycky jenom o redukce – ostnokožci (Echinodermata) jsou skupina anatomicky tak podivná, že je prakticky nemožné jim porozumět jinak než pomocí studia embryí a larev. Kořeny ostnokožců jsou patrně předkambrijské a nejstarší známé formy vypadaly ještě bizarněji než ty dnešní, zdánlivě pětipaprsčité, pocházející patrně z kambria. Za zmínku však stojí jedna z pěti dnešních skupin, okurkovití sumýši (Holothuroida). Od všech ostatních ostnokožců se liší vznikem (opět zdánlivé) sekundární „bilaterální symetrie“ – obvykle si je představujeme jako modifikované „ježovky“ (jimž jsou blízce příbuzní), protažené ve směru osy od ústního otvoru k řitnímu a následně položené na bok. To je ovšem mylná interpretace; se sumýši je ta potíž, že kromě deseti destiček v ústní oblasti nemají žádný skelet, který bychom mohli srovnávat se skeletem ostatních ostnokožců. Dnes se domníváme, že pouze ústní oblast sumýše s chapadly odpovídá celému tělu ježovky a ostatních ostnokožců – to ostatní je sumýší novotvar. Jde zřejmě o důsledek extrémního zrychlení ontogeneze, takže sumýš dospívá, aniž by vůbec dorazil ke skutečné metamorfóze. Tato drastická změna tělního plánu se odehrála patrně v siluru, ne-li dokonce v devonu, tedy z hlediska kambrijské exploze dosti nedávno (o 100 milionů let – neboli o celých „20 % geologického času“ – později).
Mohla-li takto podivná zvířata vzniknout dlouho po ukončení kambria, nebudou tělní plány objevivší se v kambriu patrně ničím zvláštním.
Tento text je úryvkem z knihy:
Jan Zrzavý, Hynek Burda, David Storch, Sabine Begallová, Stanislav Mihulka
Jak se dělá evoluce
Labyrintem evoluční biologie
Argo a Dokořán 2017
O knize na stránkách vydavatele

