 Sciencemag.cz
Sciencemag.cz
Related Articles


Základní vlastností prvních genetických molekul musela být schopnost se replikovat, tedy dělat své kopie. Zákon života je neúprosný – co se nedokáže účinně replikovat, v běhu času vymizí a informace je navždy ztracena. Pravidlo, které platí nyní a platilo i pro první replikátory.
Kopírování dávných molekul RNA určitě nebylo zcela přesné. Ba naopak, populace mírně odlišných molekul RNA mohla být materiálem pro výběr těch nejlepších v daném prostředí. Zpočátku kopírování často začínalo náhodně v libovolném místě a vznikaly neúplné kopie, zatímco úplné kopie byly v menšině. Pokud se však nějaká varianta osvědčila, bylo výhodné, aby byla zkopírována celá. Podobně jako nám příliš nevadí, když oxeroxujeme jen část stránky obsahující nesmyslný text, avšak záleží nám na tom, abychom okopírovali celou stránku, je-li obsah zajímavý.
Postupně byly zvýhodněny molekuly RNA, které zajistily replikaci celé své molekuly, například tím, že začátek kopírování nějak označily. Enzym katalyzující replikaci (pravděpodobně ribozym, tedy enzymaticky aktivní RNA) pak přesně věděl, kde má začít. Jak označit začátek replikace? Odpovědí byl zajímavý evoluční vynález – vytvoření smyčky na jednom z konců molekuly RNA. Tak bylo zajištěno zkopírování celé genetické informace. Po zkopírování však smyčka ztratila svůj smysl, mohla být dokonce svou prostorovou strukturou překážkou pro další realizaci genetické informace zkopírovaných molekul RNA. Bylo výhodné ji odstranit.
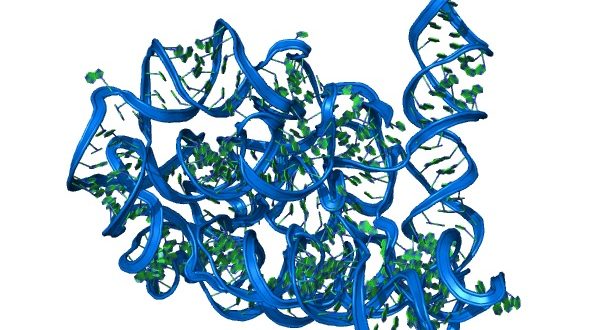
Zmíněné smyčky RNA měly další pozoruhodnou vlastnost, uměly vázat aminokyseliny. A právě této jejich vlastnosti evoluce využila. Navíc smyček bylo mnoho a mohly díky své podobnosti vzájemně interagovat. Aminokyseliny nesené na jejich koncích se tak mohly dostat do kontaktu a mohly se spojit. I takto nějak mohl vzniknout první protein. Protože řazení smyček s aminokyselinami nebylo nijak kódováno, bylo pořadí aminokyselin v takto vzniklých proteinech náhodné. Bylo jen otázkou pravděpodobnosti a selekčních tlaků, které proteiny budou mít vlastnosti výhodné pro RNA genomy.
Vidíme tedy, že z krátkých smyček, jež byly součástí delších molekul RNA, mohly vzniknout molekuly transferových RNA. Struktura jetelového listu, charakteristická pro tRNA, je odrazem jejich dávného původu. Z původní role značky určující počátek replikace primitivního RNA genomu se vyvinula dnešní úloha transferové RNA při tvorbě proteinů. Jedná se o nádherný příklad, jak evoluce pro svou tvorbu bere vše, co má po ruce. Třeba i původní genetický odpad. Předkové tRNA tak možná představovali jedny z nejstarších biomakromolekul. Pozůstatkem z dávných dob je možná i přítomnost podivných písmen genetické abecedy v molekulách tRNA – kromě bází adeninu, cytosinu, guaninu a uracilu se v tRNA nacházejí i báze inosin a hypoxantin. V tomto kontextu je zajímavé také to, že i některé dnešní mobilní genetické elementy, takzvanéskákající geny neboli transpozony, o nichž budeme vyprávět později, využívají k zahájení své replikace právě molekuly tRNA. Přesněji řečeno, tRNA slouží jako primer pro zahájení zpětné transkripce u LTR retrotranspozonů, které ve svém životním cyklu střídají fázi DNA a RNA (více viz kapitola 27).
Největší překvapení a současně podporu výše uvedených představ připravily vědcům viry. Konkrétně jeden bakteriální virus, označovaný jako bakteriofág Qβ, jehož genetická informace je uložena v molekulách RNA. Na jednom konci jeho RNA byla nalezena vlásenková struktura nápadně podobná struktuře tRNA. Tato struktura byla, podobně jako každá dnešní tRNA, zakončena trojicí písmen CCA. A co více, ukázalo se, že i v tomto případě trojice písmen CCA váže aminokyseliny, stejně jako je tomu při tvorbě proteinů, u fága Qβ však aminokyseliny hrají roli v procesu replikace.
V podobě bakteriofága Qβ máme před sebou zřejmě starobylou strukturu, jakousi molekulární fosilii přežívající z dob, kdy aminokyseliny plnily ještě jinou roli než jako stavební kameny pro syntézu proteinů. Molekuly podobné dnešním tRNA, a zejména prostorové struktury, jichž nabývaly, mohly kdysi hrát důležitou roli v nejzákladnějším procesu živých systémů, jímž je replikace. O evoluční provázanosti replikace a tvorby proteinů svědčí i fakt, že u některých virů jsou součásti replikázových komplexů proteiny podobné proteinům tvořícím ribozomy, tedy místa syntézy proteinů. Což opět naznačuje, že původní role v replikaci se přetvořila v úlohu při syntéze proteinů, přičemž u některých organismů (virů) byla dávná role dodnes zachována.
Původní molekuly tRNA byly menší, než jsou dnes, vždyť představovaly pouhou krátkou vlásenku schopnou vázat aminokyseliny. Jejich postupnou expanzí vznikly složitější molekuly tRNA, které již obsahovaly také antikodonové rameno nesoucí trojici bází, které se vážou na trojici bází kodonu nesenou molekulou mRNA. Podle této představy byla akceptorová část tRNA (nesoucí aminokyselinu) funkční již v pretranslačním světě (byla ribozymem) a je tedy starší než antikodonová smyčka, která vznikla až v souvislosti s genetickým kódem a translací. Takto zformované moderní tRNA, schopné vázat jak aminokyseliny, tak i RNA (předchůdkyni mRNA), mohly umožnit vznik delších molekul RNA. Díky nim bylo pořadí aminokyselin v prvních proteinech odrazem náhodného řazení prvních tRNA vlásenek. Selekce pak mohla vybírat první geny, kódující proteiny, jež byly nejužitečnější pro nejdůležitější funkci – replikaci RNA genomů.
Evoluce ve zkumavce
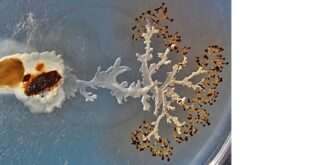
Vraťme se ještě k vlásenkové struktuře na konci molekuly RNA tvořící genom fága Qβ. Její důležitost prokázaly velice elegantní experimenty Sola Spiegelmana, jež jsou krásným příkladem selekčních procesů ve zkumavce. Měly vědcům napovědět více o počátcích života a prvních replikátorech na bázi RNA. Zejména ho zajímalo, zda se bude genom fága v průběhu experimentu nějak vyvíjet. K molekulám RNA vědci přidali enzym zajišťující kopírování RNA (RNA-polymerázu), nukleotidy potřebné pro tvorbu rostoucího řetězce RNA a trochu solí, aby zajistili vhodné prostředí.
V 60. letech 20. století, kdy Spiegelman experimenty prováděl, nebyla známa polymerázová řetězová reakce (PCR), umožňující amplifikaci v jedné zkumavce. Bylo proto nutné po každé replikaci odebrat část vzorku a přenést ho do další zkumavky, kde byla čerstvá RNA-polymeráza a nukleotidy.
Co bylo výsledkem pokusu? Protože takové zkopírování několika tisíc písmen dědičné informace nějakou dobu trvá, stalo se, že pokud tomu experimentátoři dali dostatečný čas, byla zkopírována celá RNA, tedy veškerý genom fága. Jakmile však začali dobu zkracovat nebo jinak ztěžovali podmínky kopírování, získávali stále kratší molekuly RNA. Během přenosů se genom fága neustále zmenšoval, selekce preferovala rychleji se replikující, tedy kratší genomy. Po 74 generacích (přenosech) se původní RNA genom fága Qβ o velikosti asi 4500 nukleotidů zmenšil na pouhých několik desítek nukleotidů. Co však bylo nejpozoruhodnější – vlásenka na konci byla vždy zachována, ke zkrácení docházelo v ostatních částech. Výsledný úsek odpovídal oblasti RNA, na niž se váže enzym RNA-polymeráza. Ano, tím nejdůležitějším místem genomu byla tajemná vlásenka, která se normálně nachází na konci RNA fága Qβ a určuje začátek replikace. Vše svědčilo o nepostradatelnosti vlásenky pro replikaci RNA genomu a tato struktura byla vědci označena jako „Spiegelmanovo monstrum“.
Spiegelmanův experiment tak ukázal, že i ve zkumavce, podobně jako v evoluci, je vždy zachováno to, co je momentálně důležité pro přežití.
úryvek z knihy
Eduard Kejnovský: Tajemství genů – Od vzniku života po genom člověka
2. rozšířené vydání
Academia 2026
O knize na webu vydavatele

