 Sciencemag.cz
Sciencemag.cz
Related Articles


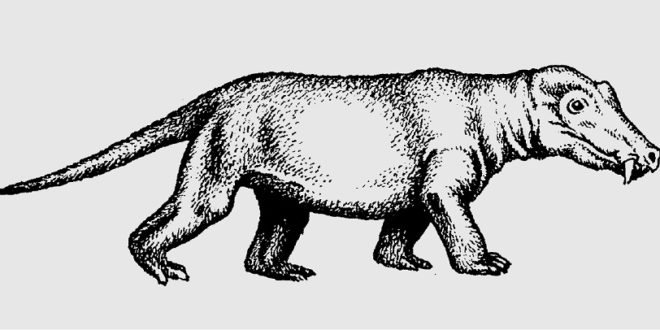
Diapsidní plazi („dinosauři“) postupně vystřídali synapsidní linii („předky savců“) v ovládání suchozemského prostředí. Jejich nástup v druhohorách je označován za triasový převrat.
Terapsidní linie se oddělila již ve středním permu a od předků podobných pelykosaurům se odlišila zejména některými znaky na lebce a čelistech (Carroll 1988, Kemp 2006). Charakteristická byla poměrně velká spánková jáma a víceméně stejně dlouhé řezáky. Evoluční přechod mezi pelykosaury a terapsidními formami je stále poněkud záhadný. Pelykosauři se až do středního permu vyskytovali především v rovníkových oblastech (v té době např. v Severní Americe a Evropě), ale fosilní nálezy terapsidů z pozdního permu jsou hlavně z lokalit ve vyšších zeměpisných šířkách (jižní Afrika, Rusko). Mezi oběma skupinami je tak patrná geografická i časová mezera. Je možné, že výměna těchto dvou velkých synapsidních radiací byla důsledkem dramatických geografických a klimatických podmínek v mladším permu a s nimi související (dosud neznámou) velkou extinkcí (Kemp 2006).
Rozhodujícím obdobím, ve kterém došlo v synapsidní linii k zásadním vývojovým změnám, byla masová extinkce na konci permu. Toto ohromné vymírání způsobilo před 250 miliony let zánik asi 70 % terestrických obratlovců a většiny suchozemských rostlin. V důsledku toho se ekosystémy a potravní řetězce zhroutily a vytvoření nových ekologických vztahů potom trvalo asi 10–30 milionů let. Rozsáhlé vymírání ukončilo převahu synapsidů a jeho důsledky uvolnily cestu k rozkvětu jiných skupin amniot, zejména archosaurních diapsidů. V terapsidní linii, která směřovala k savcům, nastalo několik důležitých změn. Hluboké změny prostředí na konci permu u synapsidních forem vytvořily selekční tlaky vedoucí k vytváření nových znaků (Carroll 1988).
Terapsidi dosáhli větší rozmanitosti tělesné stavby než pelykosauři a důležité změny nastaly v postoji těla, neboť terapsidní formy používaly výkonnější způsob kvadrupední lokomoce díky podsouvání končetin pod tělo. Původní poloha končetin ovšem zůstala v některých liniích částečně zachována, jak je patrné i u dnešních ptakořitných. Cynodontní terapsidi sice v průběhu triasu převzali úlohu dominantních predátorů od permské skupiny Gorgonopsia, ale záhy se dostali do kompetice o karnivorní niku právě s rychle se rozvíjejícími archosaury. Tato dosud málo nápadná skupina začala v triasu ekologicky i fylogeneticky nabývat převahy a diapsidní plazi postupně vystřídali synapsidní linii v ovládání suchozemského prostředí. Jejich nástup v druhohorách je označován za triasový „převrat“. Úspěch archosaurů se zdůvodňuje jejich účinnou schopností hospodařit s vodou, která mohla nabýt významu v suchém podnebí časného triasu. Archosauři (Archosauria) měli v zásadě hladký povrch těla pokrytý šupinami a produktem vylučování byla nerozpustná kyselina močová, cynodonti naproti tomu spotřebovávali vodu při rozpouštění močoviny. Výhodou mohlo být i účinnější dýchání, které podporoval vzpřímený postoj a bipední chůze archosaurů. Tato vysvětlení adaptivní převahy diapsidních archosaurů však nejsou obecně přijímána.
Dále se zdokonalovala endotermie a postupně se vyvíjelo sekundární tvrdé patro, což patrně souviselo se zvyšováním úrovně metabolických procesů. Tvrdé patro umožňovalo současně přijímat potravu i dýchat. Kostěná výztuž patra rovněž poskytovala pevný povrch, o který se mohl opírat jazyk při zpracovávání potravy a polykání. U savců patro vytvářejí dvě kosti, ale u permských synapsidů byla tato struktura budována i jinými způsoby a měla rozmanitou anatomickou stavbu.
…
Cynodonti vedli aktivní způsob života, založený na endotermní termoregulaci. Na rozvinutou schopnost produkce tepla ukazuje přítomnost chlupů. Některé formy byly asi hrabavé a žily pod zemí (např. rod Trirachodon z časného triasu) a mohla se u nich vytvářet složitější sociální struktura. Kořeny cynodontní linie sahají do prvohor, ale ze středního permu není žádný fosilní doklad, přestože v této době procházela radiací jejich sesterská skupina Therocephalia. Původní linie Cynodontia nicméně existovala více než 70 milionů let a nejstarší nálezy jsou ze svrchního permu (Charassognathidae). Z konce permu jsou potom známy již vyhraněné linie (Dviniidae, Procyonosuchidae, Galeasauridae, Thrinaxodontidae). Thrinaxodon byl archetypální časný cynodont, který také hrabal nory a mohl je sdílet s temnospondylními čtvernožci (Fernandez et al. 2013). Evoluční rozkvět cynodontů začal až po masové extinkci na hranici permu a triasu a jejich triasový fosilní záznam je lepší než permský.
Hlavní větev triasové radiace cynodontů představují Eucynodontia, kteří se dělili na převážně herbivorní Cynognathia a karnivorní Probainognathia (Abdala 2007). Při bazální divergenci karnivorních cynodontů ze skupiny Probainognathia došlo k osifikaci a uzavření frontoparietální fontanely, která je v podobné poloze jako pineální otvor. Zkostnatění fontanely ovlivňuje homeotický gen MSX2, jehož exprese souvisí také s osifikací středního ucha, vývojem mléčných žláz a mozečku (cerebellum) i udržováním vlasových (chlupových) váčků. Je pravděpodobné, že na fylogenetické úrovni těchto cynodontů se objevila zásadní mutace genu MSX2, která mohla později vést ke vzniku souvislé srsti na povrchu těla a stimulovat i další změny v linii směřující k savcům (Benoit et al. 2016). Pokročilí cynodonti nebo časní savci se museli uchýlit do prostředí s minimálním kompetičním tlakem archosaurů a později dinosaurů. Byli často býložraví (Tritylodontidae), noční a malí, existovaly však i výjimky masožravců lovících jiné obratlovce. Zvýraznila se tendence k diferenciaci zubů s precizním skusem a po masové extinkci se prosazoval liliputský efekt, projevující se miniaturizací tělesné velikosti, patrně v důsledku nedostatku zdrojů (Huttenlocker 2014) nebo díky adaptaci k efektivnímu využívání nového potravního
zdroje – terestrického hmyzu. Zmenšení tělesné velikosti a rozšíření pohybových dispozic však vedlo ke zvýšení energetických ztrát a zejména vody – tento handicap pak bylo možno kompenzovat přesunem aktivity do soumračných a nočních hodin.
Pro život v noci bylo nutné vyvinout účinnou tepelnou izolaci povrchu těla a regulovat tělesnou teplotu. Selekce podporovala také zdokonalování čichového a sluchového vnímání a urychlil se vývoj středního ucha. Nárůst velikosti čichových laloků alometricky vedl ke zvětšení mozku, přičemž větší mozek naopak vyžadoval větší přísun energie. To dále zvyšovalo energetické nároky na tepelnou izolaci těla a udržování tělesné teploty. Vedlejším důsledkem nočního života mohla být ztráta dvou ze čtyř původních opsinů čípkových fotoreceptorů sítnice. Na druhou stranu se mohla zlepšit schopnost rozeznávat barvy v přítmí.
Nejpokročilejší cynodonti jsou souborem fylogeneticky problematických skupin (čeledí), které jsou nejlépe známé z časné jury. Čeledi Tritheledontidae a Tritylodontidae byly dlouho považovány za sesterské linie savců, které různými znaky připomínaly.
Tento text je úryvkem z knihy
Miloš Macholán, Jan Zima: Systém a fylogeneze savců
Academia 2021
O knize na stránkách vydavatele

